 EasySep™小鼠TIL(CD45)正选试剂盒
EasySep™小鼠TIL(CD45)正选试剂盒





产品号 #100-0020_C
用于将人胚胎干细胞(ES)和诱导多能干细胞(iPS)来源的小胶质细胞前体分化成熟为小胶质细胞的成熟试剂盒。
若您需要咨询产品或有任何技术问题,请通过官方电话 400 885 9050 或邮箱 info.cn@stemcell.com 与我们联系。
用于将人胚胎干细胞(ES)和诱导多能干细胞(iPS)来源的小胶质细胞前体分化成熟为小胶质细胞的成熟试剂盒。
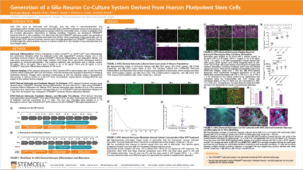
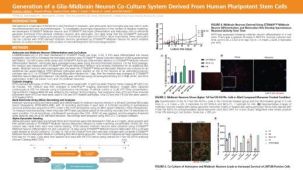
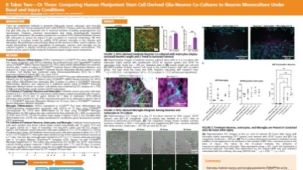
STEMdiff™ 小胶质细胞培养系统包括 STEMdiff™ 小胶质细胞分化试剂盒和 STEMdiff™ 小胶质细胞成熟试剂盒。这两种试剂盒结合使用,用于分化和成熟采用 STEMdiff™ 造血试剂盒(货号 #05310)获得的人类多能干细胞 (hPSC) 衍生小胶质细胞。
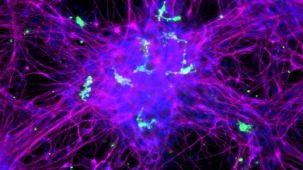
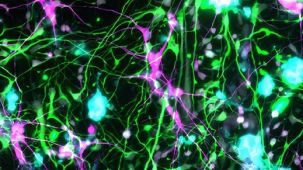
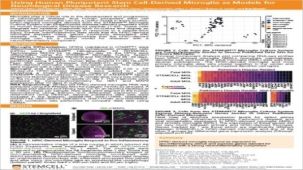
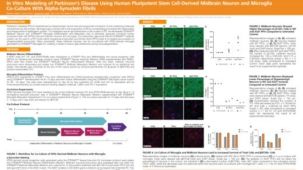
该系统基于 Mathew Blurton-Jones 实验室的方案(Abud 等,2017),生成的细胞为高纯度的小胶质细胞群(>80% CD45/CD11b 阳性,>50% TREM2 阳性小胶质细胞;形态上明显不同的单核细胞或巨噬细胞占比 <20%)。
使用该系列产品获得的细胞可作为研究神经炎症、人类神经系统发育与疾病、共培养实验以及毒性测试的多功能工具。
分类
专用培养基
细胞类型
造血细胞,PSC衍生,小胶质细胞,神经细胞,PSC衍生
种属
人
应用
细胞培养,分化
品牌
STEMdiff
研究领域
疾病建模,药物发现和毒理检测,免疫,神经科学
制剂类别
无血清
请在《产品说明书》中查找相关支持信息和使用说明,或浏览下方更多实验方案。
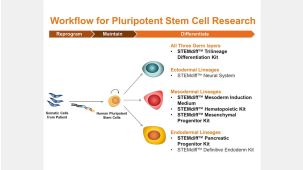
本产品专为以下研究领域设计,适用于工作流程中的高亮阶段。探索这些工作流程,了解更多我们为各研究领域提供的其他配套产品。
| 物种 | 人 |
|---|---|
| 配方 | 无血清 |

无血清神经添加物(50X)
提升神经元功能的无血清基础培养基
用于人脑类器官建立与成熟的培养基试剂盒

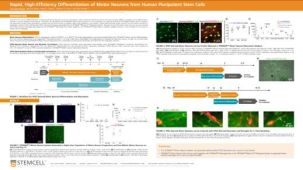
用于将人多能干细胞衍生的神经祖细胞分化为神经元前体细胞的分化试剂盒
在线联系













 沪公网安备31010102008431号
沪公网安备31010102008431号