 EasySep™小鼠TIL(CD45)正选试剂盒
EasySep™小鼠TIL(CD45)正选试剂盒





产品号 #15028_C
免疫密度负选试剂混合物
若您需要咨询产品或有任何技术问题,请通过官方电话 400 885 9050 或邮箱 info.cn@stemcell.com 与我们联系。
免疫密度负选试剂混合物
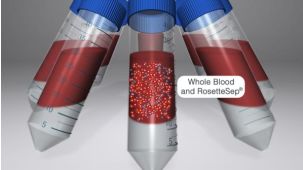
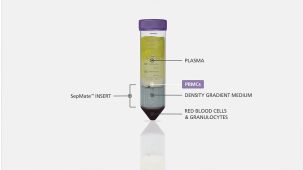
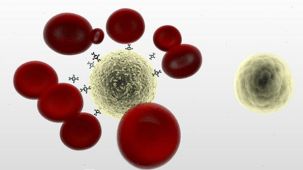
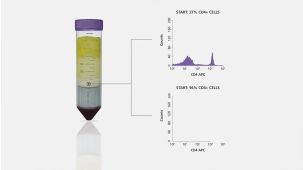
RosetteSep™人单核细胞富集抗体混合物 通过负选从全血分离单核细胞。四聚体抗体复合物可识别非单核细胞和红细胞(RBC),从而靶向去除非目的细胞。使用密度梯度离心液如Lymphoprep™(产品号 #18060)离心后 ,非目的细胞会与红细胞一起沉淀。纯化的单核细胞为血浆和密度梯度离心液的交界界面中高度富集的细胞。
分类
细胞分选试剂盒
细胞类型
单核细胞
种属
人
样本来源
白膜层、全血
分选方法
负选
应用
细胞分选
品牌
RosetteSep
研究领域
免疫
请在《产品说明书》中查找相关支持信息和使用说明,或浏览下方更多实验方案。
本产品专为以下研究领域设计,适用于工作流程中的高亮阶段。探索这些工作流程,了解更多我们为各研究领域提供的其他配套产品。
| 物种 | 人 |
|---|---|
| 样本来源 | 全血, 白膜层 |
| Selection Method | Negative |

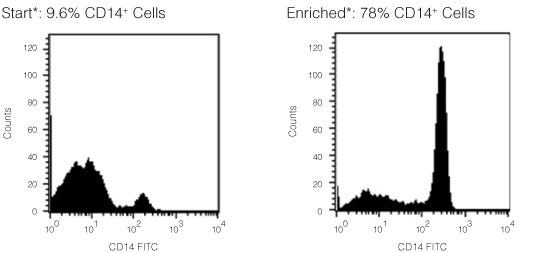
直接从全血中免疫磁珠负选人单核细胞

冻存的人原代细胞

用于体外诊断(IVD)应用的密度梯度离心管

抗人、恒河猴、食蟹猴CD14的小鼠Monoclonal IgG2a抗体

抗人、恒河猴、食蟹猴CD14的小鼠Monoclonal IgG2b抗体
在线联系





 沪公网安备31010102008431号
沪公网安备31010102008431号