 EasySep™小鼠TIL(CD45)正选试剂盒
EasySep™小鼠TIL(CD45)正选试剂盒




产品号 #05270_C
用于将人ES和iPS细胞分化为多种谱系的成分明确的无动物源培养基
若您需要咨询产品或有任何技术问题,请通过官方电话 400 885 9050 或邮箱 info.cn@stemcell.com 与我们联系。
STEMdiff™ APEL™ 2 培养基是一种成分明确、无血清、无动物源的培养基,用于人胚胎干细胞 (ES) 和诱导多能干细胞 (iPS)的分化。该培养基基于 Andrew Elefanty 博士发表的 APEL 配方,不含任何诸如无蛋白杂交瘤培养基等不明确的成分。STEMdiff™ APEL™ 2 可用于贴壁或拟胚体 (EB) 的分化方案,例如与 AggreWell™ 配合使用。该培养基可以搭配多种不同的诱导因子或细胞因子使用,以支持向外胚层、中胚层和内胚层谱系的分化。
分类
专用培养基
细胞类型
多能干细胞
种属
人
应用
细胞培养,分化
品牌
STEMdiff
研究领域
干细胞生物学
制剂类别
无动物源
请在《产品说明书》中查找相关支持信息和使用说明,或浏览下方更多实验方案。
本产品专为以下研究领域设计,适用于工作流程中的高亮阶段。探索这些工作流程,了解更多我们为各研究领域提供的其他配套产品。
| 物种 | 人 |
|---|---|
| 配方 | 无动物源性 |
高效将人多能干细胞分化为内皮细胞
用于将人 ES 细胞或 iPS 细胞分化为红系祖细胞
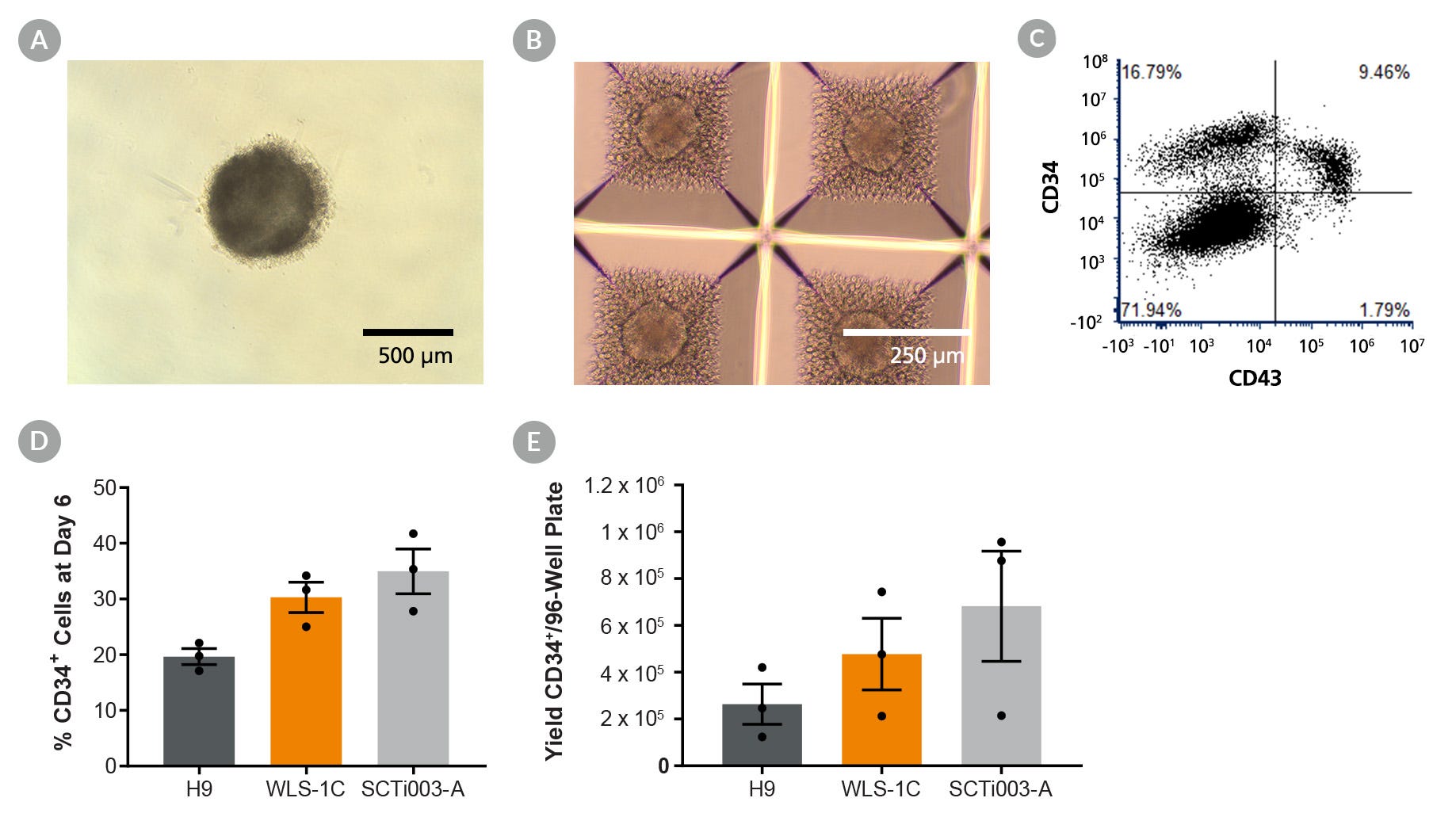
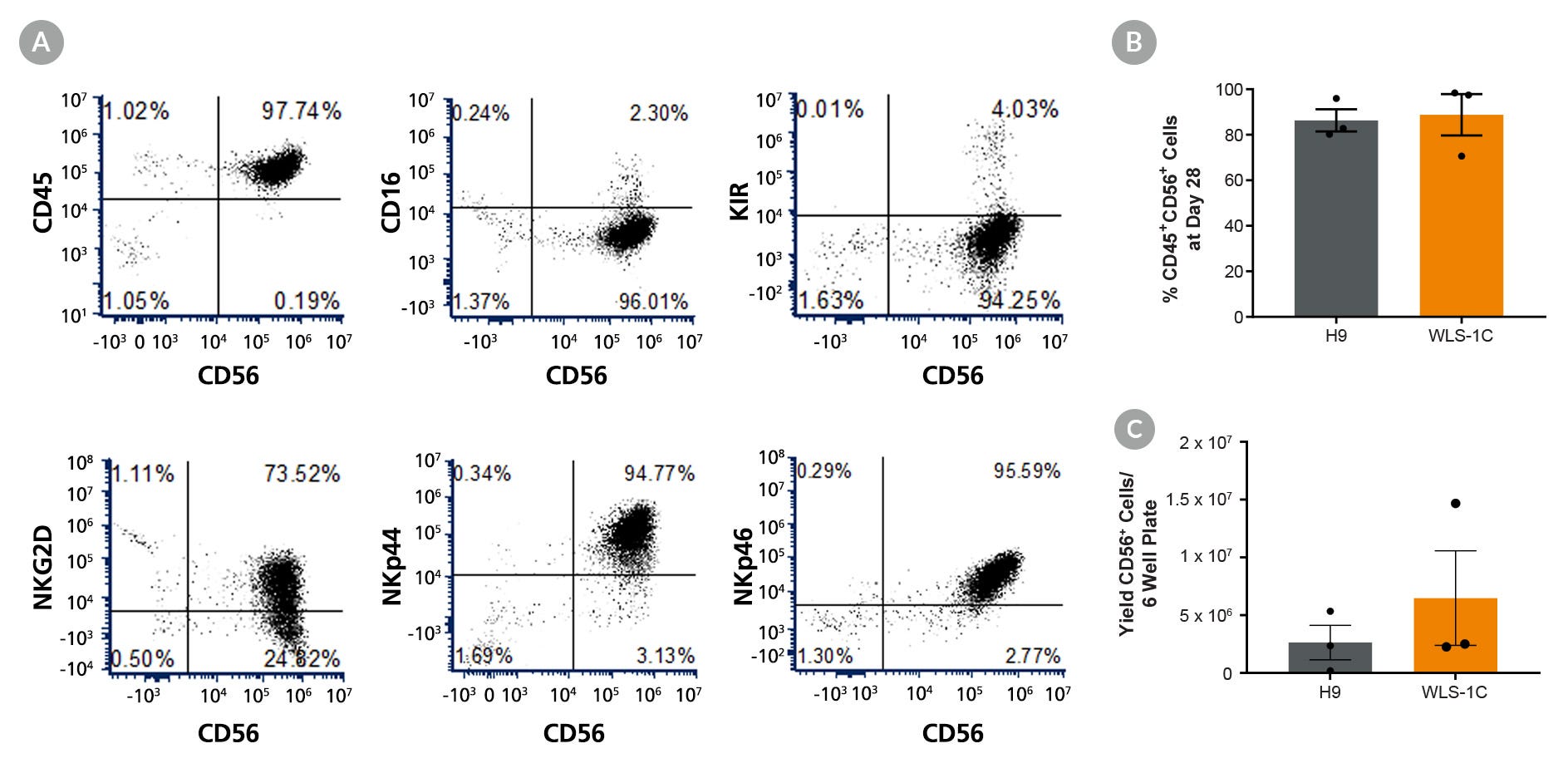
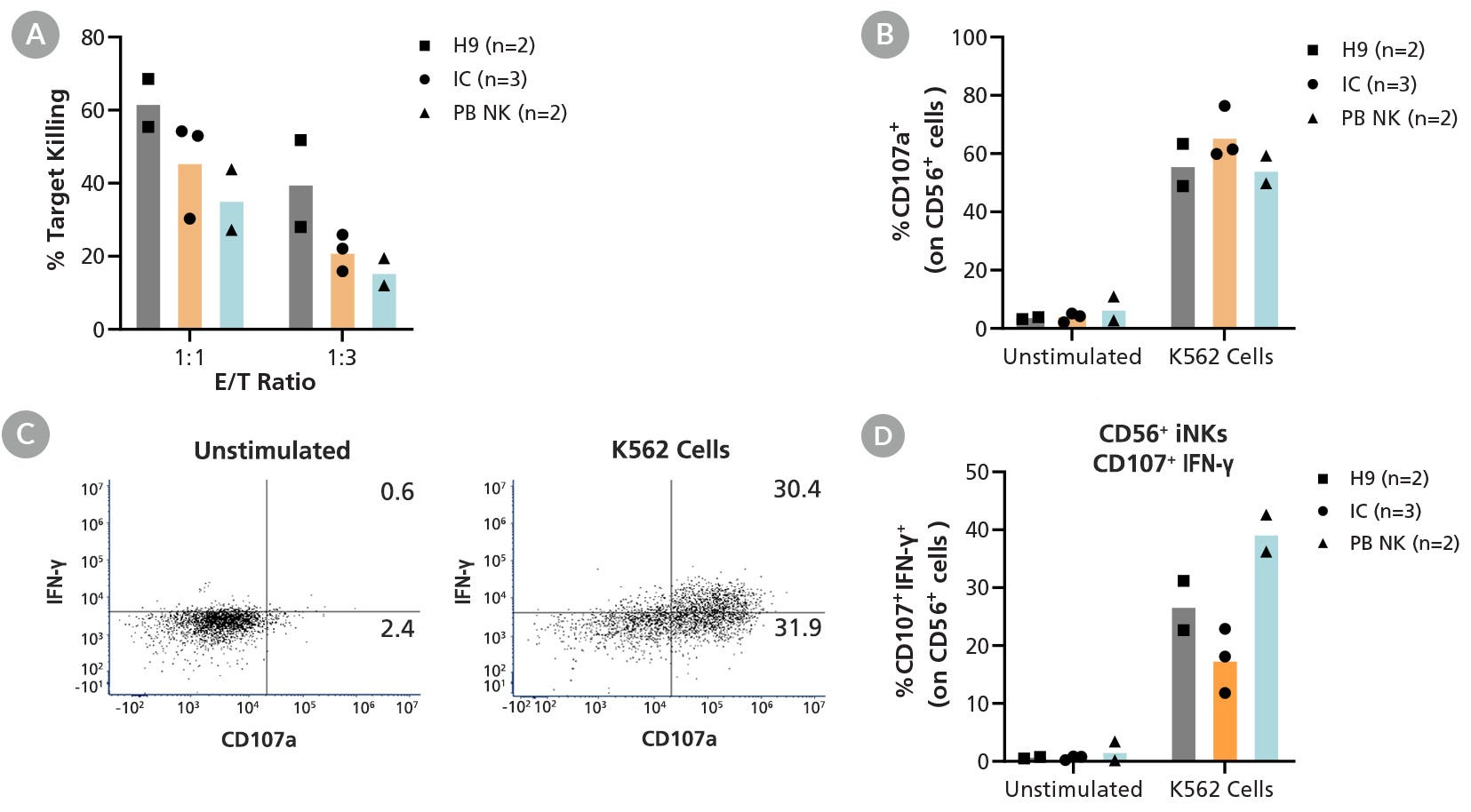
用于hPSC向NK细胞的扩增和分化

用于将人胚胎干细胞(ES细胞)或诱导多能干细胞(iPS细胞)分化为巨核细胞和血小板。
在线联系






 沪公网安备31010102008431号
沪公网安备31010102008431号