Lam AC et al. (DEC 2001)
Transfusion 41 12 1567--76
Preclinical ex vivo expansion of cord blood hematopoietic stem and progenitor cells: duration of culture; the media, serum supplements, and growth factors used; and engraftment in NOD/SCID mice.
BACKGROUND: Ex vivo expansion of cord blood (CB) hematopoietic stem and progenitor cells increases cell dose and may reduce the severity and duration of neutropenia and thrombocytopenia after transplantation. This study's purpose was to establish a clinically applicable culture system by investigating the use of cytokines,serum-free media,and autologous plasma for the expansion of CB cells and the engraftment of expanded product in nonobese diabetic/severe combined immunodeficient (NOD/SCID) mice. STUDY DESIGN AND METHODS: Enriched CB CD34+ cells were cultured in four media (Iscove's modified Dulbecco's medium with FCS,Gibco; X-Vivo-10,BioWhittaker; QBSF-60,Quality Biological; and StemSpan SFEM,Stem Cell Technologies) with four cytokine combinations (thrombopoietin [TPO],SCF,Flt-3 ligand [FL] with and without G-CSF,and/or IL-6). The effect of autologous CB plasma was also investigated. The read-out measures were evaluated on Days 8 and 12. After expansion at the optimized condition,cultured cells were transplanted into sublethally irradiated NOD/SCID mice. The engraftment of human CD45+ cells and subsets in the bone marrow,spleen,and peripheral blood was determined. RESULTS: QBSF-60 or StemSpan SFEM supported high yields of early progenitors (CD34+ cells,textlessor= 64.8-fold; CD34+CD38- cells,330-fold; CFU-granulocyte erythroid macrophage megakaryocyte [GEMM],248-fold) and CFUs of the myeloid (CFU-GM,407-fold) and erythroid (BFU/CFU-E,144-fold) lineages. The expansion of the megakaryocytic lineage was consistently higher in X-Vivo-10 (CFU-megakaryocyte,684-fold). Autologous plasma promoted colony formation but reduced CD34+ cells and CFU-GEMM. The addition of G-CSF or IL-6 improved cell yields; G-CSF was more effective for committed progenitors. Expansion products from cultures in QBSF-60 with the cytokines engrafted and differentiated into the myeloid and lymphoid lineages in NOD/SCID mice. CONCLUSION: The data supported the strategy of expansion. The optimized condition may be applicable to clinical expansion for the abrogation or reduction of posttransplant cytopenia.
View Publication
Yasui K et al. (JAN 2003)
Stem cells (Dayton,Ohio) 21 2 143--51
Differences between peripheral blood and cord blood in the kinetics of lineage-restricted hematopoietic cells: implications for delayed platelet recovery following cord blood transplantation.
Cord blood (CB) cells are a useful source of hematopoietic cells for transplantation. The hematopoietic activities of CB cells are different from those of bone marrow and peripheral blood (PB) cells. Platelet recovery is significantly slower after transplantation with CB cells than with cells from other sources. However,the cellular mechanisms underlying these differences have not been elucidated. We compared the surface marker expression profiles of PB and CB hematopoietic cells. We focused on two surface markers of hematopoietic cell immaturity,i.e.,CD34 and AC133. In addition to differences in surface marker expression,the PB and CB cells showed nonidentical differentiation pathways from AC133(+)CD34(+) (immature) hematopoietic cells to terminally differentiated cells. The majority of the AC133(+)CD34(+) PB cells initially lost AC133 expression and eventually became AC133(-)CD34(-) cells. In contrast,the AC133(+)CD34(+) CB cells did not go through the intermediate AC133(-)CD34(+) stage and lost both markers simultaneously. Meanwhile,the vast majority of megakaryocyte progenitors were of the AC133(-)CD34(+) phenotype. We conclude that the delayed recovery of platelets after CB transplantation is due to both subpopulation distribution and the process of differentiation from AC133(+)CD34(+) cells.
View Publication
Ratajczak J et al. (AUG 2011)
Leukemia 25 8 1278--85
Hematopoietic differentiation of umbilical cord blood-derived very small embryonic/epiblast-like stem cells.
A population of CD133(+)Lin(-)CD45(-) very small embryonic/epiblast-like stem cells (VSELs) has been purified by multiparameter sorting from umbilical cord blood (UCB). To speed up isolation of these cells,we employed anti-CD133-conjugated paramagnetic beads followed by staining with Aldefluor to detect aldehyde dehydrogenase (ALDH) activity; we subsequently sorted CD45(-)/GlyA(-)/CD133(+)/ALDH(high) and CD45(-)/GlyA(-)/CD133(+)/ALDH(low) cells,which are enriched for VSELs,and CD45(+)/GlyA /CD133(+)/ALDH(high) and CD45(+)/GlyA(-)/CD133(+)/ALDH(low) cells,which are enriched for hematopoietic stem/progenitor cells (HSPCs). Although freshly isolated CD45(-) VSELs did not grow hematopoietic colonies,the same cells,when activated/expanded over OP9 stromal support,acquired hematopoietic potential and grew colonies composed of CD45(+) hematopoietic cells in methylcellulose cultures. We also observed that CD45(-)/GlyA(-)/CD133(+)/ALDH(high) VSELs grew colonies earlier than CD45(-)/GlyA(-)/CD133(+)/ALDH(low) VSELs,which suggests that the latter cells need more time to acquire hematopoietic commitment. In support of this possibility,real-time polymerase chain reaction analysis confirmed that,whereas freshly isolated CD45(-)/GlyA(-)/CD133(+)/ALDH(high) VSELs express more hematopoietic transcripts (for example,c-myb),CD45(-)/GlyA(-)/CD133(+)/ALDH(low) VSELs exhibit higher levels of pluripotent stem cell markers (for example,Oct-4). More importantly,hematopoietic cells derived from VSELs that were co-cultured over OP9 support were able to establish human lympho-hematopoietic chimerism in lethally irradiated non-obese diabetic/severe combined immunodeficiency mice 4-6 weeks after transplantation. Overall,our data suggest that UCB-VSELs correspond to the most primitive population of HSPCs in UCB.
View Publication
 EasySep™小鼠TIL(CD45)正选试剂盒
EasySep™小鼠TIL(CD45)正选试剂盒




 MethoCult™ Express 含重组细胞因子的甲基纤维素培养基,用于快速进行人造血细胞集落测定
MethoCult™ Express 含重组细胞因子的甲基纤维素培养基,用于快速进行人造血细胞集落测定
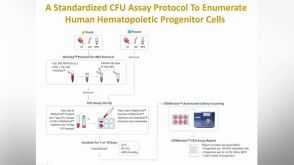
 产品手册SmartDish™ and STEMgrid™-6 Meniscus-Free Cultureware for More Accurate Counting of Hematopoietic Colonies
产品手册SmartDish™ and STEMgrid™-6 Meniscus-Free Cultureware for More Accurate Counting of Hematopoietic Colonies 产品手册STEMvision™ Automated and Standardized Counting of CFU Assays of Human Hematopoietic Cells
产品手册STEMvision™ Automated and Standardized Counting of CFU Assays of Human Hematopoietic Cells



 沪公网安备31010102008431号
沪公网安备31010102008431号